With COVID-19 on the scene, talk of other respiratory viruses has taken a backseat. But this doesn’t change the fact that they are as present as ever,…
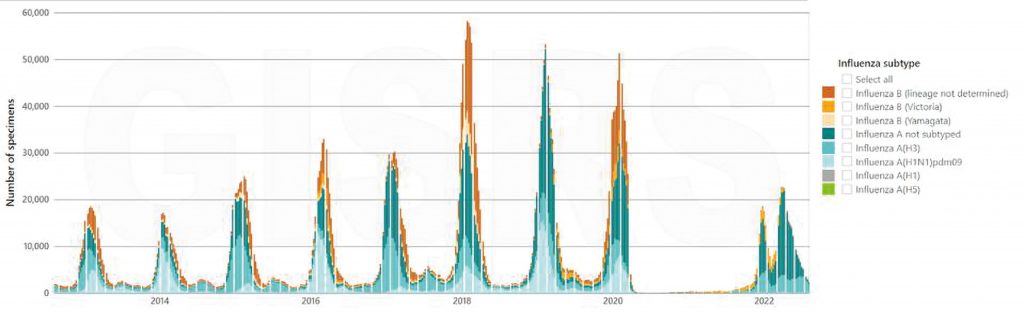
With COVID-19 on the scene, talk of other respiratory viruses has taken a backseat. But this doesn’t change the fact that they are as present as ever, lurking in the background. Influenza is one such virus, causing an estimated 1 billion cases globally every year—of these, upwards of 500,000 end up being fatal. Although lockdowns and social distancing efforts aimed at curbing the spread of COVID-19 also helped put a dent in seasonal influenza epidemics, the flu is now returning with full force (Figure 1).
Figure 1. Number of specimens positive for influenza by subtype globally, from August 2012 to August 2022. Source: FLUNET (www.who.int/toolkits/flunet). global influenza surveillance and response system (GISRS). Data generated on 08/10/2022
Recently there has been some success on the road to developing a much sought-after broadly neutralizing vaccine against all influenza A viruses. If successful, it may act as a precursor to a truly universal flu vaccine, one that protects against all types and subtypes of the virus. The breakthrough may also provide a blueprint for developing a COVID-19 vaccine that retains its efficacy in the face of new variants.
Understanding influenza
There are seven types of influenza, four of which can infect mammals: A, B, C, and D. Influenza A and B are the most common, driving yearly seasonal flu epidemics. Influenza C infections in humans are rare and are thought to cause relatively mild disease. Influenza D mainly infects cattle with no recorded human infections to date.
On the whole, influenza A viruses are considered the biggest threat. All of the major influenza pandemics of the last century have been caused by influenza A viruses, the most notable being the Spanish flu pandemic of 1918 which led to the death of roughly 50 million people, one-tenth of the entire global population at the time.
Influenza A viruses are also particularly adept at evading our immune system; they have a high rate of genetic mutation, resulting in a process known as antigenic variation. This is what allows the virus to reinfect us again every season, it is what makes the virus volatile and capable of causing the occasional pandemic, and it is what makes it so difficult for us to develop long-lasting vaccines. To help explain antigenic variation, it’s worth quickly looking at the structure of the virus (Figure 2).
Figure 2. Schematic of influenza A virion structure. [Timonina/Shutterstock.com]
First, all influenza viruses have single-stranded, negative-sense RNA. This is the genetic material used by the virus to replicate. In the case of influenza A, the genome is broken into eight separate segments. Surrounding and protecting these RNA strands is the viral envelope. The envelope contains two key surface proteins hemagglutinin (HA) and neuraminidase (NA)—the former helps the virus enter our cells, and the latter helps newly-formed viral particles leave the cell.
There are 18 different subtypes of the hemagglutinin surface protein, aptly named H1 through H18. These are split into two groups, organized according to their evolutionary history. Crucially, immunity against one group does not confer immunity against the other group. Any universal influenza A vaccine has to account for this fact. Neuraminidase, in turn, has 11 different subtypes—N1 through N11.
It is on the basis of these surface protein subtypes that influenza A viruses are classified. For example, the viral subtype that caused the 1918 Spanish flu had an H1 hemagglutinin protein and a N1 neuraminidase protein, making it an influenza A (H1N1) virus. The influenza virus that caused the 1968 pandemic, in contrast, had a different HA subtype, H3, and a different NA subtype as well, N2. As a result, it is classified as influenza A (H3N2). These are the two most common subtypes of influenza A, responsible for the majority of yearly infections.
Influenza A subtypes can be broken down even further into strains. This is determined by the sum of their genetic material, known as the genome. Two viruses may have the same broad hemagglutinin and neuraminidase subtypes, but have subtle differences throughout their genome. For example, the H1N1 subtype was responsible for both the 1918 and 2009 influenza pandemics, but in each case it was a different strain of the virus. (Figure 3).
Figure 3. A rough timeline of the most recent major influenza pandemics. Genetic mutations characterizing each pandemic strain are labeled in the center of the viral particles; each line represents an RNA segment, which encodes different proteins carried by the virus. From: “The Evolution and Future of Influenza Pandemic Preparedness” Harrington et al. 2021
So, how does all of this relate to antigenic variation and why does it make it difficult for us to develop a universal influenza A vaccine?
The HA and NA surface proteins are recognized by our immune system as foreign substances—known as antigens. Detection of antigens in our body leads to the activation of various immune responses aimed at clearing the threat. One kind of immune response involves training B cells to produce antibodies that specifically target the antigens in question. In a sense, our B cells develop a type of “immune memory” towards the virus. Doing so allows our immune system to react more quickly and effectively in case of reinfection, blocking the virus before it has the chance to get us sick.
As hinted at above, influenza A viruses are notorious for their ability to mutate. This happens when the virus makes small “mistakes” while copying its genetic information during replication. Over time, these small mutations to the genome can accumulate, changing the structure of various parts of the virus, including HA and NA. If it changes enough, the virus will be able to evade our immune system; our B cells will no longer be able to recognize the antigens, nullifying what they may have learned during a prior infection. This is a type of antigenic variation called “drift”, and it is the reason we can repeatedly become sick with the flu.
Vaccination against influenza works by exposing our immune system to killed or weakened forms of the virus. In the same way a natural infection would, this helps our immune system build up a memory of the viral antigens and teaches it to keep an eye out for them in the future. But like the antibodies produced after natural infection, those produced in response to vaccination are also rendered useless over time by antigenic drift; as before, the antigens become too different to be recognized, forcing B cells to rebuild their immune memory all over again. This is why scientists have to develop a new flu vaccine every year, so that our immune system can build up its memory based on the most recent circulating influenza antigens.
There’s also another, more drastic kind of antigenic variation called antigenic “shift”. Whereas drift happens over an extended period of time, via the accumulation of small changes, antigenic shift can lead to substantial changes very quickly. How? Usually when a cell is infected by an influenza virus, eight gene segments come in, these eight segments are copied and packaged into a newly formed viral particle, and the new particle then leaves the cell to go infect other cells. But sometimes a single cell—whether human or avian or porcine—can become infected by two different influenza A viruses at the same time. In such cases, the two viruses can end up “exchanging” RNA segments with one another. This is called reassortment, a process that leads to the creation of a third, novel virus that’s distinct from both of the parent strains. Often this happens when an influenza A virus jumps between different animal hosts. For example, the 1957 influenza pandemic was caused by a novel subtype, H2N2, composed partly of genes from the 1918 strain and partly from new genes gained while infecting birds (Figure 3).
Such sudden jumps in genetic code can lead to influenza viruses that look very different to those usually circulating, rendering the surface proteins unrecognizable to our immune system. Once again, our immune system is confronted with what is essentially an entirely new threat and has to build its immune memory from the ground up. Whereas antigenic drift is responsible for yearly seasonal flu outbreaks, antigenic shift is often responsible for new influenza pandemics. Any universal influenza A vaccine has to successfully account for both antigenic drift and antigenic shift.
The seasonal approach
In an attempt to circumvent the issue of antigenic variation, vaccine manufacturers update the flu shot each year based on the latest circulating influenza strains. In the United States, all influenza vaccines are “quadrivalent”, meaning they contain four different influenza viruses. This is done to broaden protection against the various influenza subtypes and lineages known to drive seasonal outbreaks: influenza A (H1N1), influenza A (H3N2), influenza B/Victoria, and influenza B/Yamagata. Quadrivalent vaccines will also protect against any other influenza viruses that are antigenically similar.
Unfortunately, vaccines produced in this way are nowhere near as effective as we might hope. At best, they protect 60% of people from illness, but this number can, and often does, drop much lower. For the influenza A (H3N2) subtype, vaccine effectiveness hovers around 33%. So where are we going wrong?
Missing the target: Egg-based vaccines
Selection of candidate vaccine viruses (CVVs) is only one part of the equation, growing them is another. This is no simple feat considering they need to be available in bulk, enough to make millions of vaccines. For the past 70 years, the majority of manufacturers have turned to chicken eggs in order to achieve the necessary growth (Figure 4). The candidate vaccine viruses are injected into fertilized hen’s eggs and left to incubate for a few days. During this period, the viruses are able to replicate. The fluid in the eggs is then extracted and the viruses are “killed” (inactivated). Finally, the antigen of choice—usually the hemagglutinin surface protein—is isolated from the killed viruses and purified, making it ready for use in vaccines. Even now, most flu vaccines continue to be egg-based.
Figure 4. An overview of the steps involved in producing egg-based vaccines. [Esco Vaccixcell]
In response to these issues, manufacturers have tried to develop new production methods that avoid using chicken eggs to culture candidate viruses. Cell-based and recombinant vaccines aim to resolve the issues on the production side of things. The former by skipping the need for eggs, and by extension, the threat of egg-adapted changes. The latter by skipping the need for eggs as well as cutting down the time it takes to produce the vaccines, reducing the risk of escape mutants.
However, there is a paucity of comparative research on these new production methods, making it difficult to draw any firm conclusions, but early findings suggest recombinant vaccines may be more effective than traditional egg-based and cell-based vaccines, including improved antibody production.
Possibility of mRNA influenza vaccines?
Messenger RNA (mRNA) technology could also help solve the issues associated with slow production. Unlike traditional flu vaccines, mRNA vaccines do not depend on the cultivation of viruses. Instead, they are based on the synthesis of carefully designed mRNA. The process is more akin to how a pharmaceutical drug like aspirin is produced than it is to how current flu shots are produced.
As soon as researchers have a complete sequence of a virus’ genome, they can isolate the section they want to use for vaccination—usually, the gene corresponding to the surface protein the virus uses to bind and enter our cells—and artificially synthesize it in the lab. The gene can then be inserted into a small, circular piece of DNA known as a plasmid. Plasmids are easy to replicate, allowing researchers to produce large amounts of the target viral gene at once. Finally, the plasmids are put into a reactor with a molecule that transcribes the viral RNA into mRNA. Any remaining plasmid material is broken down and the mRNA put through a series of purifiers before being packaged into the protective lipid shell. The whole process takes a matter of weeks rather than months. Or maybe even less: in 2013, a team of scientists at Novartis generated an mRNA vaccine candidate in the span of eight days.
Unlike COVID-19 outbreaks, which are usually driven by one dominant circulating variant, seasonal flu outbreaks are characterized by multiple cocirculating strains. These strains are derived from the four main influenza viruses: influenza A(H3N2), influenza A(H1N1), and two influenza B viruses. To account for differences in the hemagglutinin surface protein between strains, traditional flu shots are “quadrivalent”, meaning they contain an inactivated version of each of the four viruses. But a synthesized mRNA sequence can only make one particular protein; an effective mRNA influenza vaccine would need four different sequences to make four different HA proteins. A 2020 study led by researchers at the Icahn School of Medicine at Mount Sinai suggests this shouldn’t be an issue. The team of scientists manufactured an mRNA vaccine against an influenza subtype that encoded four different proteins at once. They have since repeated the experiment for two additional influenza subtypes, and they plan to test a combined vaccine that encodes for a total of 10 to 12 different antigens across subtypes. This is part of a larger movement towards mRNA flu shots.
What remains unclear is whether mRNA technology by itself could address a major issue plaguing current influenza vaccines: waning immunity. A 2018 meta-analysis of influenza vaccine durability found that protection can vanish as early as 90 days post vaccination. Recombinant and inactivated vaccines both suffer from this issue, even though they trigger immune responses in different ways. If the COVID-19 mRNA vaccines are anything to go by, waning immunity will likely continue to be a factor even if influenza vaccines made the switch to mRNA technology.
Nasal influenza vaccines
There has been tremendous excitement around mucosal vaccines, with many in the field pinning their hopes on long-term protection from infection as well as severe disease. I do not share the same unbridled enthusiasm that nasal vaccines will provide long-lasting protection from infection and protection from all variants for several reasons.
The first nasal flu vaccine, FluMist, was approved by the U.S. Food and Drug Administration (FDA) in 2012. By 2016 it had been pulled off the market, not to reappear until 2018. The reason? Decreased vaccine effectiveness. Combined data from 2013 to 2016 indicates that nasal spray vaccine effectiveness was roughly 26% in children between the ages of 12 and 17. This is compared to 51% effectiveness for inactivated vaccines, administered intramuscularly.
Although these low numbers were brushed off as being a result of poorly matched vaccine virus strains, there may also be other reasons the nasal spray influenza vaccine hasn’t quite lived up to expectations.
For one, IgA is typically quite short-lived; it is produced only for a brief period after exposure. Any barrier immunity that the nasal spray vaccine does offer us will likely fade as soon as IgA numbers drop. And durability is already a point of concern with traditional, inactivated flu vaccines, which depend on the longer-lived circulating IgG class of antibodies for protection from infection. So, we cannot expect it to be all that durable.
Another worry is that the nasal spray vaccine may be cleared by our immune system before it has the chance to complete its job. Adults who have previously been exposed to the flu—and that means more or less every adult—will still have at least partial mucosal immunity against the influenza viruses. The attenuated, or “weakened”, viruses used in the nasal spray vaccine might simply be getting neutralized before they can infect nasal cells, preventing our B cells from updating sIgA to match the viruses circulating that season.
On a more theoretical note, one might also question the belief that vaccine-induced mucosal immunity can provide lasting protection if natural infection, through the same path of entry, fails to do so. Many people are infected by influenza on a seasonal basis, with waning immunity and viral mutation leading to renewed vulnerability year after year. This happens even though they would have built up some degree of mucosal immunity during prior infection. If prior infection through the nose doesn’t protect us from reinfection, why would a nasal vaccine?
Vaccine platform vs. vaccine antigens
How the antigen is delivered is one side of the coin, what antigen gets chosen in the first place is the other side. Delivery platforms continue to evolve, yet progress in antigen selection has been lagging. To optimize our influenza vaccines, we need both.
Targeting conserved regions: HA stalk and M2e
Whether egg-based, cell-based, recombinant or otherwise, most of the currently available flu shots use the hemagglutinin (HA) surface protein as their antigen of choice. This makes sense; influenza viruses depend on the hemagglutinin protein to gain entry into our cells. Block the protein, and you have a chance at blocking infection. The issue with using hemagglutinin as the primary antigen is that it is extremely mutable. The structure of the protein can change to avoid previously established immunity, including B cell-derived antibodies. Thus, the seasonal flu cycle and yearly flu shots.
The universal vaccine approach aims to change things. Instead of using the head of the hemagglutinin protein as the antigen, researchers working towards an universal flu vaccine look for viral proteins, or specific protein regions, that are highly conserved across strains and across time—essentially, antigens that only very rarely mutate. If successful, universal vaccines could provide a much broader immunity, eliminating the need for new vaccines every year.
So far, there have been two promising contenders: the stalk portion of the hemagglutinin protein and, for influenza A viruses, the extracellular region of the Matrix-2 protein (M2e).
The hemagglutinin surface protein can be separated into two regions, one being the highly mutable head (HA1) and the other being the stable stem (HA2) (Figure 5). The head region contains the receptor binding domain (RBD) which the influenza viruses use to bind to our cells. Using this area as the reference antigen creates a highly specific immune response, with antibodies that can block the RBD and, if present in sufficient numbers, may prevent infection. But the specificity comes at the cost of breadth. Even a single mutation can throw off our immune system, rendering the antibodies practically ineffective.
Figure 5. “The influenza virus with surface proteins hemagglutinin (HA) and neuraminidase (NA) is shown in the left panel. The middle panel shows the X-ray structure of the HA trimeric glycoprotein. The right panel shows the phylogenetic tree of HA, group 1 (colored blue) and group 2 (colored red). The subtypes that have been confirmed in humans are boxed.” From: “Antibody cross-reactivity to hemagglutinin protein antigens demonstrates feasibility for development of a “universal” influenza a synthetic peptide vaccine” Jiang et al. 2015
Where the HA head domain is used to bind to our cells, the stem region is used for fusion—the process by which the virus “injects” its genetic material into the host cell cytoplasm, kicking off viral replication. Briefly, once a viral particle has bound to a host cell it is “ingested” by a section of the cell membrane that then buds off to form a membrane “bubble”. The resulting bubble is called a vesicle, and foreign substances internalized in this way are usually promptly digested and disposed of by the cell. But influenza viruses are crafty and have developed a way to use the acid that normally helps our cells digest such vesicle contents to instead free up the RNA inside the viral particle. This is where the stem portion of the hemagglutinin protein comes into play. Once the RNA has been freed up, the HA stem is used as a lever of sorts to pull the viral membrane into the cellular membrane, finally creating a “tear” that lets the genetic material spill out into the cytoplasm. Once there, the virus can begin replicating before spreading to other cells.
A vaccine that targets the HA stem and blocks it from initiating fusion would interrupt the replication process, preventing further infection and stopping the virus in its tracks. Indeed, researchers have already developed vaccines based on “headless” HA proteins as antigens, but results so far have been slightly underwhelming; although the protection the vaccines have provided has been fairly broad, it has not been very strong. Mice inoculated with vaccines based on HA stalk as an antigen still lose noticeable amounts of body weight, a common symptom of illness. This has been particularly true for influenza A viruses with group 2 HA proteins (Figure 5), which also happen to be those causing the majority of seasonal infections.
In response, attention has also been directed towards the matrix-2 (M2) protein (Figure 6). This protein spans the membrane of the influenza viral particle and functions to internalize the acid I mentioned above. Without the M2 protein, the viral RNA cannot be untethered from its membrane and replication is once again impaired. Vaccines that target the extracellular domain of M2 (M2e) have fared better than those that target the HA head alone.
Figure 6. Viral particle of influenza with its membrane proteins: hemagglutinin (HA), neuraminidase (NA), matrix protein 2 (M2); and associated proteins: matrix protein 1 (M1), ribonucleoprotein (RNP). A) A model of the viral particle. B) Viral budding. Credit: “infuenza M2 proton channels” Pielak et al. 2011
More recently, efforts have been made to merge HA stem and M2e into one single molecule, to boost immunogenicity. Linking them chemically has yielded promising results, with higher protective potency than HA stem or M2e vaccines alone.
Design of the antigen: Perfecting the “headless” HA protein
Interest in the stem portion of HA has its roots in the discovery of human broadly neutralizing antibodies (bnAbs) that target the region. This discovery revitalized hopes of developing universal flu vaccines by giving researchers an entry point for the production of a universal antigen. Since the full HA protein only rarely elicits broadly neutralizing antibodies, various approaches have emerged to expose and properly present the stem domain to our immune system. One of the more successful strategies involves knocking off the HA head domain while ensuring structural stability of the leftover HA stem.
But this is easier said than done. The head of the HA protein stabilizes the prefusion protein resulting in a tightly coiled triplex of three proteins. Broadly neutralizing monoclonal antibodies only recognize this tightly coiled triplex and are lost once the fusion process commences or the head of the HA protein is removed.
To perfect the hemagglutinin portion of their antigen, Subbiah and colleagues drew upon previous research by Impagliazzo et al. and Yassine et al. Both groups managed to engineer stable, headless HA stem antigens—called “mini-HAs”—that exhibited structural and antibody binding properties similar to full-length HA. Antibodies produced in response to these mini-HAs managed to protect against a wide range of HAs in mice and nonhuman primates.
Figure 7. “(A) The mini-HA design strategy is described in five stages, each characterized by specific modifications leading to selection of the best construct for each stage. Key modifications for each stage are schematically depicted and color-coded. (B) Ribbon mini-HA model (from PDB ID 1RD8) with color-coded modifications (per stage, as in Fig. 1A) and putative N-glycosylation sites (CG1-3, CG7; taupe).” From: “A stable trimeric influenza hemagglutinin stem as a broadly protective immunogen” Impagliazzo et al. 2015
Figure 7 outlines the various stages of Impagliazzo et al.’s design, culminating in the creation of a stable, headless “mini-HA” stem that retains all the relevant neutralizing binding sites. Figure 8 outlines the design stages of Yassine et al, who achieved great minimization of the HA protein, ultimately succeeding in having the stem domain contribute 94% of the total surface area of their antigen.
Figure 8. (A) Progressive removal of the HA head domain to expose as much of the HA stem domain as possible without losing structural stability. (B) Percentage of HA stem contribution to total antigen surface area: full HA protein (left), Gen4 of modified “headless” HA antigen (center), and Gen6 of modified headless HA antigen. From: “hemagglutinin-stem nanoparticles generate heterosubtypic influenza protection” Yassine et al. 2015
Forming a stable trimer and preventing protein aggregation
Influenza hemagglutinin is naturally a trimeric molecule, meaning it is composed of three distinct hemagglutinin proteins bound together. In order to accurately mimic this three-part structure, Subbiah et al. excluded the C-terminus of the HA stem portion and instead replaced it with the foldon sequence of a virus that infects bacteria.
Figure 9. (A) A trimeric fiber loses its structure and breaks down into unfolded monomers, leading to irreversible protein aggregation. (B) The addition of the foldon sequence allows monomers to return to their trimeric structure, preventing loss of structure and subsequent protein aggregation. From: “foldon-guided self-assembly of ultra-stable protein fibers” Bhardwaj et al. 2009
When added to the end of a monomer that has a propensity to dissociate, the foldon sequence stabilizes it and allows it to form a trimer (Figure 9). Failure to do so may lead the headless HA stem antigen to lose its structure, and by extension its epitope sites. If our antibodies cannot bind to the stem, they cannot learn about its structure and thus cannot take on the specific shape needed to protect us from future reinfection.
The researchers also added specific point mutations along HA1 head and HA2 stalk to prevent protein aggregation issues. All of the mutations were in regions outside of those recognized by broadly neutralizing antibodies. Each one of the mutations was in a long helical region with a tendency to self-aggregate. By changing the amino acids in these stretches, they substantially reduced non-specific aggregation and instead favored the stable form of the protein.
In addition to this, they mutated a cysteine residue and replaced it with a serine residue, again strengthening the stability of the protein and minimizing the chance of any unwanted disulfide bonds from forming.
Addition of M2e
To broaden the neutralization of their antigen and increase its potency, the researchers added in two identical segments of the M2 channel protein. The segments they chose correspond to a 23 amino acid-long sequence that encodes the epitopes for the extracellular region of the protein. This area protrudes from the viral membrane, allowing it to be bound by neutralizing antibodies.
Crucially, M2e has previously been shown to confer cross-group immunity. Influenza A viruses are split up into two groups according to phylogenetic differences in their HA protein. Usually immunity against group 1 HAs (H1, H2, H5, H6, H8, H9, H11, H12, H13, H16, H17, H18) does not confer immunity against group 2 HAs (H3, H4, H7, H10, H14, H15). This means that infection by an influenza A/(H3N2) virus, for example, generally does not confer protection against a strain of influenza A/(H1N1).
Finally, Subbiah et al. connected the various domains of the M2e-H3 construct using flexible linkers, which are small amino acids that function like bungee cords; they keep the proteins tethered together but are flexible enough not to interfere with the folding process.
Figure 10. M2e-H3 stalk vaccine construct with flexible and soluble linker sequences (AAAGGAA; GGGGS; GSA; GSAGSA; QGTGG).” From: “A chimeric thermostable M2e and H3 stalk-based universal influenza a virus vaccine” Subbiah et al. 2022Figure 11. A schematic diagram of the H3 HA protein, derived from the H3N2 influenza virus. (Above) A schematic of the chimeric M2e-H3 stalk protein, with stabilizing point mutations marked in violet. From: Subbiah et al. 2022
The full genetic sequence of the M2e-H3 antigen can be seen in Figure 10, and the structure of the antigen—as compared to wild type hemagglutinin—can be seen in Figure 11.
Production and purification of the antigen
For ease of purification of the antigen, the researchers added a histidine tag (6xHis) to the N-terminal of the M2e-H3 stem domain. A histidine tag is made up of a DNA sequence that encodes six to nine histidine residues. These histidine residues bind to several different types of immobilized metal ions. So, once a protein has been “His-tagged” it can be easily identified and separated from other proteins by exposure to such metal ions. The histidine tag can then be removed from the protein or it can be retained, either way it does not affect the antigenicity of the protein.
Next, Subbiah et al. synthesized a codon-optimized gene encoding the M2e-H3 stem protein. This gene is then inserted into escherichia coli (E. coli) for quick and easy mass production.
Does it retain antigenicity?
After extraction and purification, Subbiah et al. began testing the M2e-H3 stalk molecule for its ability to be recognized by antibodies, known as antigenicity. They exposed the chimeric molecule to antibodies known to target the stem portion of the hemagglutinin protein as well as antibodies known to target M2e; in both cases they witnessed high reactivity. The scientists recorded similar results from antisera—blood samples from mice that had previously been infected with an influenza virus and had built up a corresponding antibody response. Importantly, the construct retained its antigenicity when exposed to influenza A viruses belonging to different HA groups. And, even after storage at 122 degrees Fahrenheit for 11 days, the antigen continued to elicit strong antibody reactivity, suggesting solid thermostability.
Animal models
Having confirmed that the M2E-H3 stem protein can be recognized and bound by antibodies, Subbiah and colleagues moved on to vaccination trials. Often, vaccines based on viral proteins instead of inactivated or attenuated viruses require an adjuvant to help stimulate a stronger immune response, and by extension, better protective efficacy. For their experiments, the researchers used a particularly strong adjuvant similar to the AS01 liposome adjuvant licensed for use in herpes Zoster vaccination. This is a double-edged sword: the adjuvant ensures a quick immune response, but because of its strength, it also comes at the cost of side effects. Although the side effects are not much cause for concern—headache, fever, chills, and so on— they are uncomfortable all the same. Using a different delivery platform, like mRNA technology, may help circumvent these issues in the future.
The adjuvanted M2e-H3 stalk vaccine was administered to mice as two doses, a primary vaccination followed by a boost two weeks later. Following the first dose, there was a noticeable increase in antibodies specific to M2e and HA stem regions. Addition of the boost further increased antibody titers 10 fold. Antisera built up in response to M2e-H3 exposure managed to protect against both group 1 and group 2 influenza A viruses. Subbiah et al. suspect this might be explained by antibody-dependent cellular cytotoxicity (ADCC), a mechanism of immune defense whereby effector cells recognize antibodies stuck to antigens and actively break down the foreign threat. Their suspicions were proven right by western blotting, which showed that M2e antibodies, in particular, raised ADCC activity 20 fold.
Figure 12. Protective efficacy comparison of M2e-H3 stalk and M2e only vaccines after A/HK/1968 H3N2 (12xLD50, 1.4 × 102 EID50) virus challenge. From: Subbiah et al. 2022
To compare protection against an M2e only vaccine, the group of researchers designed another chimera protein combining M2e fused with the receptor binding domain (RBD) of SARS-CoV-2 spike protein. When exposed to a lethal dose of influenza A/HK/H3N2 virus, all of the mice inoculated with M2e-H3 stem survived, suffering only moderate body weight loss of around 10%. On the other hand, none of the mice inoculated with the M2e-only vaccine survived the infection (Figure 12).
Along with a robust antibody response, M2e-H3 stalk vaccination also triggered a strong T cell response. Where antibodies can bind to pathogens to prevent infection, T cells are in charge of getting rid of already-infected cells, curbing the spread of the virus. Indeed, the vaccine significantly reduced the amount of virus in the lungs of infected mice within a span of six days. Depleting T cell levels in vaccinated mice before exposing them to a lethal challenge with influenza A H7N9 produced twice as much weight loss (12%) than in mice that had not been depleted of their T cells.
The work by Subbiah et al. acts as a clear proof of concept: using M2e and HA stalk regions as antigens can stimulate broad neutralizing immunity. Crucially, this immunity should hold up to new strains, as it targets those viral regions that only very rarely mutate. Although their work focused specifically on influenza A viruses, there is no reason to think that, given enough time and research, we won’t be able to produce a truly universal influenza vaccine.
William R. Haseltine, PhD, is chair and president of the think tank ACCESS Health International, a former Harvard Medical School and School of Public Health professor and founder of the university’s cancer and HIV/AIDS research departments. He is also the founder of more than a dozen biotechnology companies, including Human Genome Sciences.
"I Can't Even Save": Americans Are Getting Absolutely Crushed Under Enormous Debt Load
While Joe Biden insists that Americans are doing great - suggesting in his State of the Union Address last week that "our economy is the envy of the world," Americans are being absolutely crushed by inflation (which the Biden admin blames on 'shrinkflation' and 'corporate greed'), andof course - crippling debt.
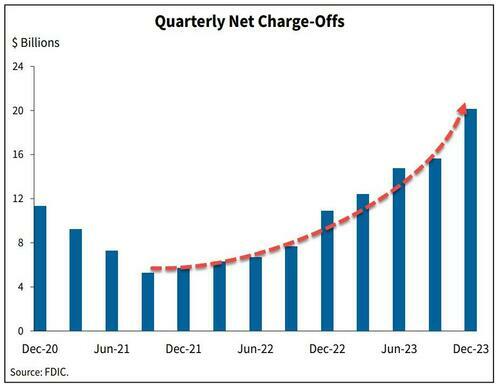
The signs are obvious. Last week we noted that banks' charge-offs are accelerating, and are now above pre-pandemic levels.
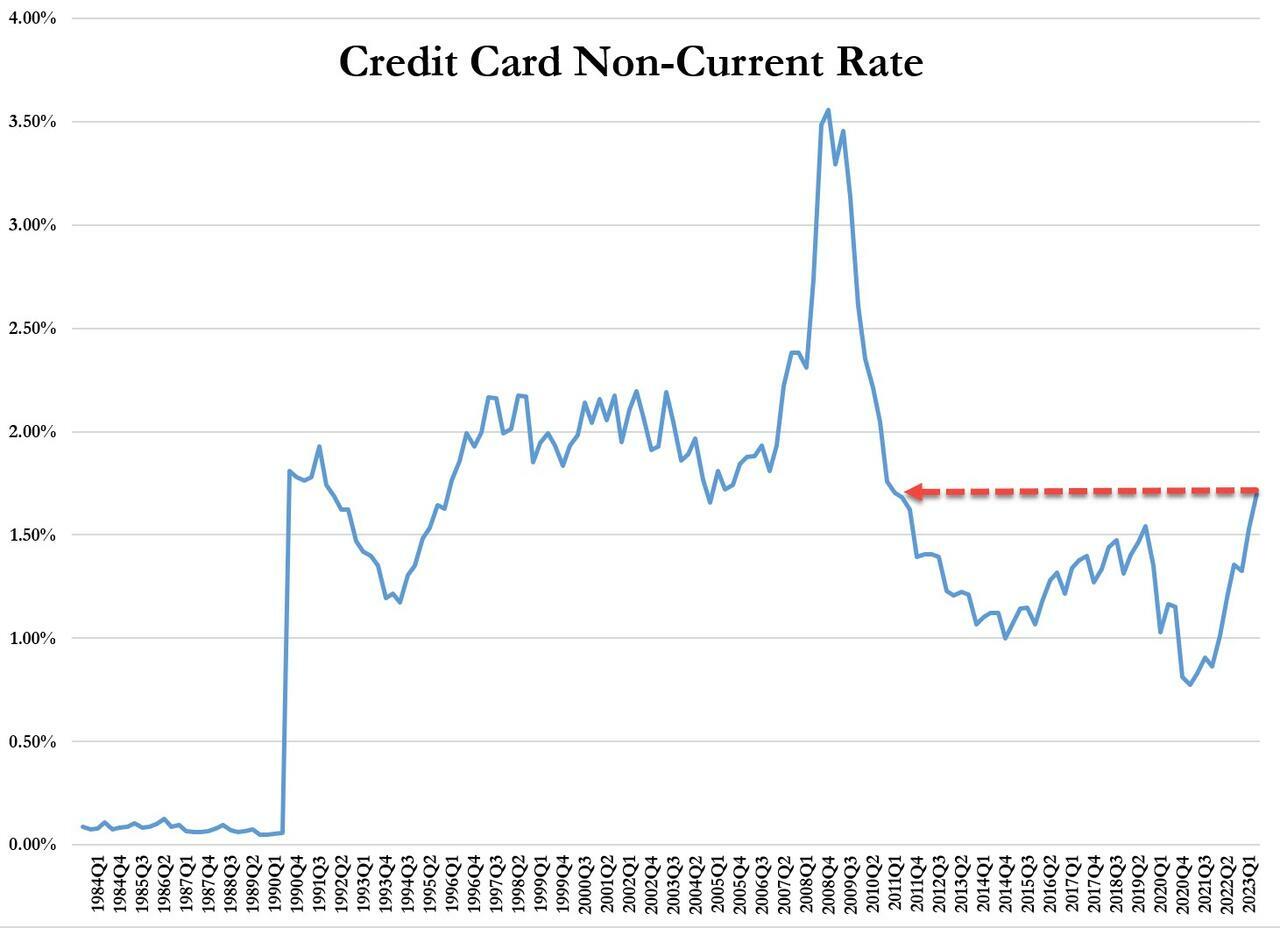
...and leading this increase are credit card loans - with delinquencies that haven't been this high since Q3 2011.
On top of that, while credit cards and nonfarm, nonresidential commercial real estate loans drove the quarterly increase in the noncurrent rate, residential mortgages drove the quarterly increase in the share of loans 30-89 days past due.
And while Biden and crew can spin all they want, an average of polls from RealClear Politics shows that just 40% of people approve of Biden's handling of the economy.
Crushed
On Friday, Bloomberg dug deeper into the effects of Biden's "envious" economy on Americans - specifically, how massive debt loads (credit cards and auto loans especially) are absolutely crushing people.
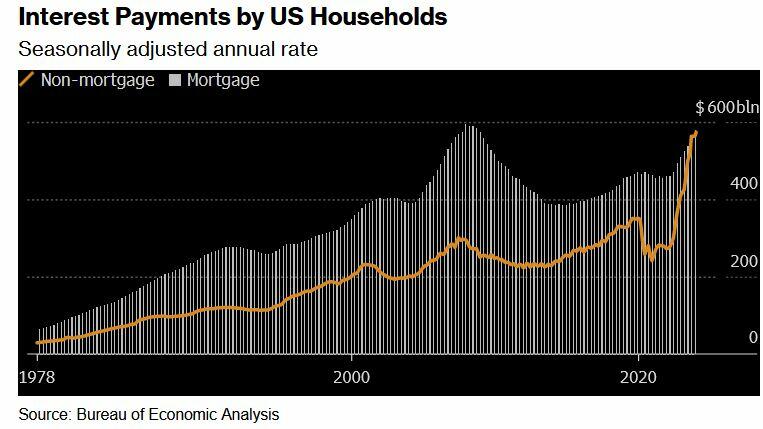
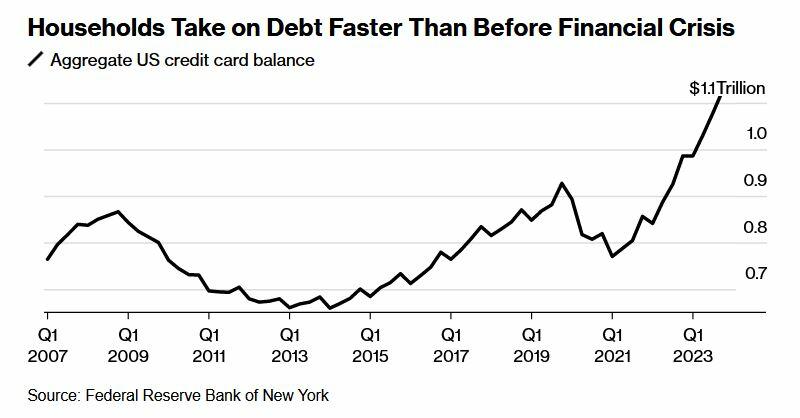
Two years after the Federal Reserve began hiking interest rates to tame prices, delinquency rates on credit cards and auto loans are the highest in more than a decade. For the first time on record, interest payments on those and other non-mortgage debts are as big a financial burden for US households as mortgage interest payments.
According to the report, this presents a difficult reality for millions of consumers who drive the US economy - "The era of high borrowing costs — however necessary to slow price increases — has a sting of its own that many families may feel for years to come, especially the ones that haven’t locked in cheap home loans."
The Fed, meanwhile, doesn't appear poised to cut rates until later this year.
According to a February paper from IMF and Harvard, the recent high cost of borrowing - something which isn't reflected in inflation figures, is at the heart of lackluster consumer sentiment despite inflation having moderated and a job market which has recovered (thanks to job gains almost entirely enjoyed by immigrants).
In short, the debt burden has made life under President Biden a constant struggle throughout America.
"I’m making the most money I've ever made, and I’m still living paycheck to paycheck," 40-year-old Denver resident Nikki Cimino told Bloomberg. Cimino is carrying a monthly mortgage of $1,650, and has $4,000 in credit card debt following a 2020 divorce.
"There's this wild disconnect between what people are experiencing and what economists are experiencing."
CBS: Do you attribute the inflation crisis to the pandemic or Biden?
WISCONSIN VOTER: "It's been YEARS now since the pandemic — I'm not buying that anymore. At first I did; I'm not buying that anymore because yogurt is STILL going up in price!" pic.twitter.com/apahb65scB
What's more, according to Wells Fargo, families have taken on debt at a comparatively fast rate - no doubt to sustain the same lifestyle as low rates and pandemic-era stimmies provided. In fact, it only took four years for households to set a record new debt level after paying down borrowings in 2021 when interest rates were near zero.
Meanwhile, that increased debt load is exacerbated by credit card interest rates that have climbed to a record 22%, according to the Fed.
[P]art of the reason some Americans were able to take on a substantial load of non-mortgage debt is because they’d locked in home loans at ultra-low rates, leaving room on their balance sheets for other types of borrowing. The effective rate of interest on US mortgage debt was just 3.8% at the end of last year.
Yet the loans and interest payments can be a significant strain that shapes families’ spending choices. -Bloomberg
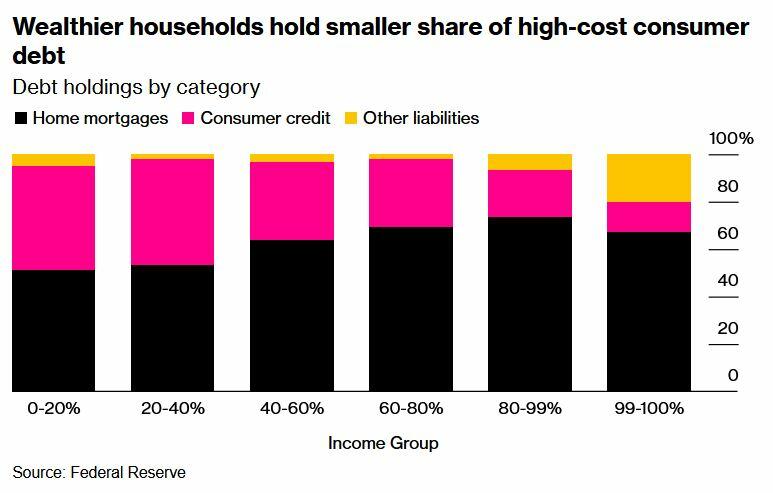
And of course, the highest-interest debt (credit cards) is hurting lower-income households the most, as tends to be the case.
The lowest earners also understandably had the biggest increase in credit card delinquencies.
"Many consumers are levered to the hilt — maxed out on debt and barely keeping their heads above water," Allan Schweitzer, a portfolio manager at credit-focused investment firm Beach Point Capital Management told Bloomberg. "They can dog paddle, if you will, but any uptick in unemployment or worsening of the economy could drive a pretty significant spike in defaults."
"We had more money when Trump was president," said Denise Nierzwicki, 69. She and her 72-year-old husband Paul have around $20,000 in debt spread across multiple cards - all of which have interest rates above 20%.
During the pandemic, Denise lost her job and a business deal for a bar they owned in their hometown of Lexington, Kentucky. While they applied for Social Security to ease the pain, Denise is now working 50 hours a week at a restaurant. Despite this, they're barely scraping enough money together to service their debt.
The couple blames Biden for what they see as a gloomy economy and plans to vote for the Republican candidate in November. Denise routinely voted for Democrats up until about 2010, when she grew dissatisfied with Barack Obama’s economic stances, she said. Now, she supports Donald Trump because he lowered taxes and because of his policies on immigration. -Bloomberg
Meanwhile there's student loans - which are not able to be discharged in bankruptcy.
"I can't even save, I don't have a savings account," said 29-year-old in Columbus, Ohio resident Brittany Walling - who has around $80,000 in federal student loans, $20,000 in private debt from her undergraduate and graduate degrees, and $6,000 in credit card debt she accumulated over a six-month stretch in 2022 while she was unemployed.
"I just know that a lot of people are struggling, and things need to change," she told the outlet.
The only silver lining of note, according to Bloomberg, is that broad wage gains resulting in large paychecks has made it easier for people to throw money at credit card bills.
Yet, according to Wells Fargo economist Shannon Grein, "As rates rose in 2023, we avoided a slowdown due to spending that was very much tied to easy access to credit ... Now, credit has become harder to come by and more expensive."
According to Grein, the change has posed "a significant headwind to consumption."
Then there's the election
"Maybe the Fed is done hiking, but as long as rates stay on hold, you still have a passive tightening effect flowing down to the consumer and being exerted on the economy," she continued. "Those household dynamics are going to be a factor in the election this year."
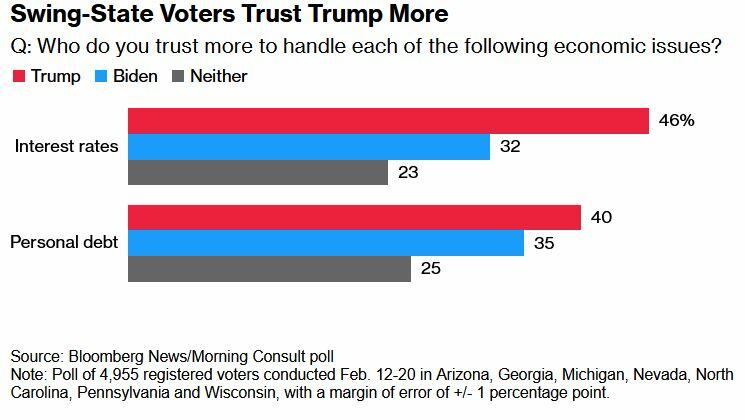
Meanwhile, swing-state voters in a February Bloomberg/Morning Consult poll said they trust Trump more than Biden on interest rates and personal debt.
Reverberations
These 'headwinds' have M3 Partners' Moshin Meghji concerned.
"Any tightening there immediately hits the top line of companies," he said, noting that for heavily indebted companies that took on debt during years of easy borrowing, "there's no easy fix."
Sylvester researchers, collaborators call for greater investment in bereavement care
MIAMI, FLORIDA (March 15, 2024) – The public health toll from bereavement is well-documented in the medical literature, with bereaved persons at greater…
MIAMI, FLORIDA (March 15, 2024) – The public health toll from bereavement is well-documented in the medical literature, with bereaved persons at greater risk for many adverse outcomes, including mental health challenges, decreased quality of life, health care neglect, cancer, heart disease, suicide, and death. Now, in a paper published in The Lancet Public Health, researchers sound a clarion call for greater investment, at both the community and institutional level, in establishing support for grief-related suffering.
Credit: Photo courtesy of Memorial Sloan Kettering Comprehensive Cancer Center
MIAMI, FLORIDA (March 15, 2024) – The public health toll from bereavement is well-documented in the medical literature, with bereaved persons at greater risk for many adverse outcomes, including mental health challenges, decreased quality of life, health care neglect, cancer, heart disease, suicide, and death. Now, in a paper published in The Lancet Public Health, researchers sound a clarion call for greater investment, at both the community and institutional level, in establishing support for grief-related suffering.
The authors emphasized that increased mortality worldwide caused by the COVID-19 pandemic, suicide, drug overdose, homicide, armed conflict, and terrorism have accelerated the urgency for national- and global-level frameworks to strengthen the provision of sustainable and accessible bereavement care. Unfortunately, current national and global investment in bereavement support services is woefully inadequate to address this growing public health crisis, said researchers with Sylvester Comprehensive Cancer Center at the University of Miami Miller School of Medicine and collaborating organizations.
They proposed a model for transitional care that involves firmly establishing bereavement support services within healthcare organizations to ensure continuity of family-centered care while bolstering community-based support through development of “compassionate communities” and a grief-informed workforce. The model highlights the responsibility of the health system to build bridges to the community that can help grievers feel held as they transition.
The Center for the Advancement of Bereavement Care at Sylvester is advocating for precisely this model of transitional care. Wendy G. Lichtenthal, PhD, FT, FAPOS, who is Founding Director of the new Center and associate professor of public health sciences at the Miller School, noted, “We need a paradigm shift in how healthcare professionals, institutions, and systems view bereavement care. Sylvester is leading the way by investing in the establishment of this Center, which is the first to focus on bringing the transitional bereavement care model to life.”
What further distinguishes the Center is its roots in bereavement science, advancing care approaches that are both grounded in research and community-engaged.
The authors focused on palliative care, which strives to provide a holistic approach to minimize suffering for seriously ill patients and their families, as one area where improvements are critically needed. They referenced groundbreaking reports of the Lancet Commissions on the value of global access to palliative care and pain relief that highlighted the “undeniable need for improved bereavement care delivery infrastructure.” One of those reports acknowledged that bereavement has been overlooked and called for reprioritizing social determinants of death, dying, and grief.
“Palliative care should culminate with bereavement care, both in theory and in practice,” explained Lichtenthal, who is the article’s corresponding author. “Yet, bereavement care often is under-resourced and beset with access inequities.”
Transitional bereavement care model
So, how do health systems and communities prioritize bereavement services to ensure that no bereaved individual goes without needed support? The transitional bereavement care model offers a roadmap.
“We must reposition bereavement care from an afterthought to a public health priority. Transitional bereavement care is necessary to bridge the gap in offerings between healthcare organizations and community-based bereavement services,” Lichtenthal said. “Our model calls for health systems to shore up the quality and availability of their offerings, but also recognizes that resources for bereavement care within a given healthcare institution are finite, emphasizing the need to help build communities’ capacity to support grievers.”
Key to the model, she added, is the bolstering of community-based support through development of “compassionate communities” and “upskilling” of professional services to assist those with more substantial bereavement-support needs.
The model contains these pillars:
Preventive bereavement care –healthcare teams engage in bereavement-conscious practices, and compassionate communities are mindful of the emotional and practical needs of dying patients’ families.
Ownership of bereavement care – institutions provide bereavement education for staff, risk screenings for families, outreach and counseling or grief support. Communities establish bereavement centers and “champions” to provide bereavement care at workplaces, schools, places of worship or care facilities.
Resource allocation for bereavement care – dedicated personnel offer universal outreach, and bereaved stakeholders provide input to identify community barriers and needed resources.
Upskilling of support providers – Bereavement education is integrated into training programs for health professionals, and institutions offer dedicated grief specialists. Communities have trained, accessible bereavement specialists who provide support and are educated in how to best support bereaved individuals, increasing their grief literacy.
Evidence-based care – bereavement care is evidence-based and features effective grief assessments, interventions, and training programs. Compassionate communities remain mindful of bereavement care needs.
Lichtenthal said the new Center will strive to materialize these pillars and aims to serve as a global model for other health organizations. She hopes the paper’s recommendations “will cultivate a bereavement-conscious and grief-informed workforce as well as grief-literate, compassionate communities and health systems that prioritize bereavement as a vital part of ethical healthcare.”
“This paper is calling for healthcare institutions to respond to their duty to care for the family beyond patients’ deaths. By investing in the creation of the Center for the Advancement of Bereavement Care, Sylvester is answering this call,” Lichtenthal said.
Follow @SylvesterCancer on X for the latest news on Sylvester’s research and care.
# # #
Article Title: Investing in bereavement care as a public health priority
DOI: 10.1016/S2468-2667(24)00030-6
Authors: The complete list of authors is included in the paper.
Funding: The authors received funding from the National Cancer Institute (P30 CA240139 Nimer) and P30 CA008748 Vickers).
Disclosures: The authors declared no competing interests.
# # #
Journal
The Lancet Public Health
DOI
10.1016/S2468-2667(24)00030-6
Article Title
Investing in bereavement care as a public health priority
Artificial intelligence (AI) cannot distinguish fact from fiction. It also isn’t creative or can create novel content but repeats, repackages, and reformulates what has already been said (but perhaps in new ways).
I am sure someone will disagree with the latter, perhaps pointing to the fact that AI can clearly generate, for example, new songs and lyrics. I agree with this, but it misses the point. AI produces a “new” song lyric only by drawing from the data of previous song lyrics and then uses that information (the inductively uncovered patterns in it) to generate what to us appears to be a new song (and may very well be one). However, there is no artistry in it, no creativity. It’s only a structural rehashing of what exists.
Of course, we can debate to what extent humans can think truly novel thoughts and whether human learning may be based solely or primarily on mimicry. However, even if we would—for the sake of argument—agree that all we know and do is mere reproduction, humans have limited capacity to remember exactly and will make errors. We also fill in gaps with what subjectively (not objectively) makes sense to us (Rorschach test, anyone?). Even in this very limited scenario, which I disagree with, humans generate novelty beyond what AI is able to do.
Both the inability to distinguish fact from fiction and the inductive tether to existent data patterns are problems that can be alleviated programmatically—but are open for manipulation.
Manipulation and Propaganda
When Google launched its Gemini AI in February, it immediately became clear that the AI had a woke agenda. Among other things, the AI pushed woke diversity ideals into every conceivable response and, among other things, refused to show images of white people (including when asked to produce images of the Founding Fathers).
Tech guru and Silicon Valley investor Marc Andreessen summarized it on X (formerly Twitter): “I know it’s hard to believe, but Big Tech AI generates the output it does because it is precisely executing the specific ideological, radical, biased agenda of its creators. The apparently bizarre output is 100% intended. It is working as designed.”
There is indeed a design to these AIs beyond the basic categorization and generation engines. The responses are not perfectly inductive or generative. In part, this is necessary in order to make the AI useful: filters and rules are applied to make sure that the responses that the AI generates are appropriate, fit with user expectations, and are accurate and respectful. Given the legal situation, creators of AI must also make sure that the AI does not, for example, violate intellectual property laws or engage in hate speech. AI is also designed (directed) so that it does not go haywire or offend its users (remember Tay?).
However, because such filters are applied and the “behavior” of the AI is already directed, it is easy to take it a little further. After all, when is a response too offensive versus offensive but within the limits of allowable discourse? It is a fine and difficult line that must be specified programmatically.
It also opens the possibility for steering the generated responses beyond mere quality assurance. With filters already in place, it is easy to make the AI make statements of a specific type or that nudges the user in a certain direction (in terms of selected facts, interpretations, and worldviews). It can also be used to give the AI an agenda, as Andreessen suggests, such as making it relentlessly woke.
Thus, AI can be used as an effective propaganda tool, which both the corporations creating them and the governments and agencies regulating them have recognized.
Misinformation and Error
States have long refused to admit that they benefit from and use propaganda to steer and control their subjects. This is in part because they want to maintain a veneer of legitimacy as democratic governments that govern based on (rather than shape) people’s opinions. Propaganda has a bad ring to it; it’s a means of control.
However, the state’s enemies—both domestic and foreign—are said to understand the power of propaganda and do not hesitate to use it to cause chaos in our otherwise untainted democratic society. The government must save us from such manipulation, they claim. Of course, rarely does it stop at mere defense. We saw this clearly during the covid pandemic, in which the government together with social media companies in effect outlawed expressing opinions that were not the official line (see Murthy v. Missouri).
AI is just as easy to manipulate for propaganda purposes as social media algorithms but with the added bonus that it isn’t only people’s opinions and that users tend to trust that what the AI reports is true. As we saw in the previous article on the AI revolution, this is not a valid assumption, but it is nevertheless a widely held view.
If the AI then can be instructed to not comment on certain things that the creators (or regulators) do not want people to see or learn, then it is effectively “memory holed.” This type of “unwanted” information will not spread as people will not be exposed to it—such as showing only diverse representations of the Founding Fathers (as Google’s Gemini) or presenting, for example, only Keynesian macroeconomic truths to make it appear like there is no other perspective. People don’t know what they don’t know.
Of course, nothing is to say that what is presented to the user is true. In fact, the AI itself cannot distinguish fact from truth but only generates responses according to direction and only based on whatever the AI has been fed. This leaves plenty of scope for the misrepresentation of the truth and can make the world believe outright lies. AI, therefore, can easily be used to impose control, whether it is upon a state, the subjects under its rule, or even a foreign power.
The Real Threat of AI
What, then, is the real threat of AI? As we saw in the first article, large language models will not (cannot) evolve into artificial general intelligence as there is nothing about inductive sifting through large troves of (humanly) created information that will give rise to consciousness. To be frank, we haven’t even figured out what consciousness is, so to think that we will create it (or that it will somehow emerge from algorithms discovering statistical language correlations in existing texts) is quite hyperbolic. Artificial general intelligence is still hypothetical.
As we saw in the second article, there is also no economic threat from AI. It will not make humans economically superfluous and cause mass unemployment. AI is productive capital, which therefore has value to the extent that it serves consumers by contributing to the satisfaction of their wants. Misused AI is as valuable as a misused factory—it will tend to its scrap value. However, this doesn’t mean that AI will have no impact on the economy. It will, and already has, but it is not as big in the short-term as some fear, and it is likely bigger in the long-term than we expect.
No, the real threat is AI’s impact on information. This is in part because induction is an inappropriate source of knowledge—truth and fact are not a matter of frequency or statistical probabilities. The evidence and theories of Nicolaus Copernicus and Galileo Galilei would get weeded out as improbable (false) by an AI trained on all the (best and brightest) writings on geocentrism at the time. There is no progress and no learning of new truths if we trust only historical theories and presentations of fact.
However, this problem can probably be overcome by clever programming (meaning implementing rules—and fact-based limitations—to the induction problem), at least to some extent. The greater problem is the corruption of what AI presents: the misinformation, disinformation, and malinformation that its creators and administrators, as well as governments and pressure groups, direct it to create as a means of controlling or steering public opinion or knowledge.
This is the real danger that the now-famous open letter, signed by Elon Musk, Steve Wozniak, and others, pointed to:
“Should we let machines flood our information channels with propaganda and untruth? Should we automate away all the jobs, including the fulfilling ones? Should we develop nonhuman minds that might eventually outnumber, outsmart, obsolete and replace us? Should we risk loss of control of our civilization?”
Other than the economically illiterate reference to “automat[ing] away all the jobs,” the warning is well-taken. AI will not Terminator-like start to hate us and attempt to exterminate mankind. It will not make us all into biological batteries, as in The Matrix. However, it will—especially when corrupted—misinform and mislead us, create chaos, and potentially make our lives “solitary, poor, nasty, brutish and short.”
We use cookies on our website to give you the most relevant experience by remembering your preferences and repeat visits. By clicking “Accept”, you consent to the use of ALL the cookies.
This website uses cookies to improve your experience while you navigate through the website. Out of these, the cookies that are categorized as necessary are stored on your browser as they are essential for the working of basic functionalities of the website. We also use third-party cookies that help us analyze and understand how you use this website. These cookies will be stored in your browser only with your consent. You also have the option to opt-out of these cookies. But opting out of some of these cookies may affect your browsing experience.
Necessary cookies are absolutely essential for the website to function properly. These cookies ensure basic functionalities and security features of the website, anonymously.
Cookie
Duration
Description
cookielawinfo-checbox-analytics
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Analytics".
cookielawinfo-checbox-functional
11 months
The cookie is set by GDPR cookie consent to record the user consent for the cookies in the category "Functional".
cookielawinfo-checbox-others
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Other.
cookielawinfo-checkbox-necessary
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookies is used to store the user consent for the cookies in the category "Necessary".
cookielawinfo-checkbox-performance
11 months
This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Performance".
viewed_cookie_policy
11 months
The cookie is set by the GDPR Cookie Consent plugin and is used to store whether or not user has consented to the use of cookies. It does not store any personal data.
Functional cookies help to perform certain functionalities like sharing the content of the website on social media platforms, collect feedbacks, and other third-party features.
Performance cookies are used to understand and analyze the key performance indexes of the website which helps in delivering a better user experience for the visitors.
Analytical cookies are used to understand how visitors interact with the website. These cookies help provide information on metrics the number of visitors, bounce rate, traffic source, etc.
Advertisement cookies are used to provide visitors with relevant ads and marketing campaigns. These cookies track visitors across websites and collect information to provide customized ads.













































{kind=link}
{kind=link}